

タンパク質は、人体の約5分の1を構成する重要な物質だ。ところが、「タンパク質(eiwit:蘭)」という名称がオランダの科学者ムルダーの論文で1838年に使われてから、すでに2世紀以上が経つにもかかわらず、タンパク質の全貌は未だに解明されていない。
いま分かっているのは、タンパク質を構成するアミノ酸が、多数連結して立体構造をとり、生体内で常に揺れ動きながら特定の機能を発揮することまでだ。だが、これまでの構造解析法(X線結晶構造解析やクライオ電子顕微鏡観測)で得られる構造は、ダイナミックに動き回るタンパク質の一瞬を捉えたスナップショットでしかない。タンパク質は生体内で、実際にどのように動いているのか。その謎に挑むのが、京都大学 複合原子力科学研究所の杉山正明教授だ。教授らのグループは、中性子を活用した独自の手法を用いて、タンパク質の動態観察に挑み、生体内の環境に近い溶液中で、タンパク質の構造を正確に評価する解析法の開発を目指している。
「誰も見たことのない世界を、何とかしてこの目で見たい」
研究は、こうした純粋な知的好奇心から始まることが多い。杉山教授の研究も、「生体内で複雑に動くことで、さまざまな機能を発現するタンパク質の姿を生のまま見たい」という途方もない願望からスタートした。この願望がどれほど非現実的であるかは、タンパク質の動きを考えると理解できる。
タンパク質は、いくつものアミノ酸がつながって折り畳まれて立体構造をとる。その生成過程では、まずアミノ酸が鎖状に複数つながり、タンパク質の元となる「ポリペプチド」ができる。次にポリペプチドが適切な状態に折り畳まれ、特定の立体構造となる。これが、「ドメイン」と呼ばれるタンパク質の構成単位だ。続いて、ドメインが複数連結し、ようやく1つのタンパク質が完成する。完成したタンパク質に他のタンパク質が結合し、新たな機能を発現する「会合体」となることもある。
図1に示すゾーンⅣ、つまり会合体ぐらいの大きさであれば、光学的なイメージを使ってモノ自体を観察できる。けれども、これ以下のサイズの実態を知るためには分光法を使い、そのエネルギー変化から類推することになる。その場合ゾーンⅠのアミノ酸は、サイズは小さいもののエネルギー変化が大きく激しく動いているため、その姿を分光法で捉えやすくなる。
ところがゾーンⅡからゾーンⅢのあたりは、サイズはnm(ナノメートル)スケールにとどまるうえ、エネルギーレベルもmeVやμeVと低いため、動きを捉えるのが極めて難しい。
このゾーンⅡとゾーンⅢを捉えるために、杉山教授はエネルギー分解能がほぼ最高レベルにある中性子スピンエコー法を使う。これにより測定したデータをコンピュータでシミュレーションし検証していく。
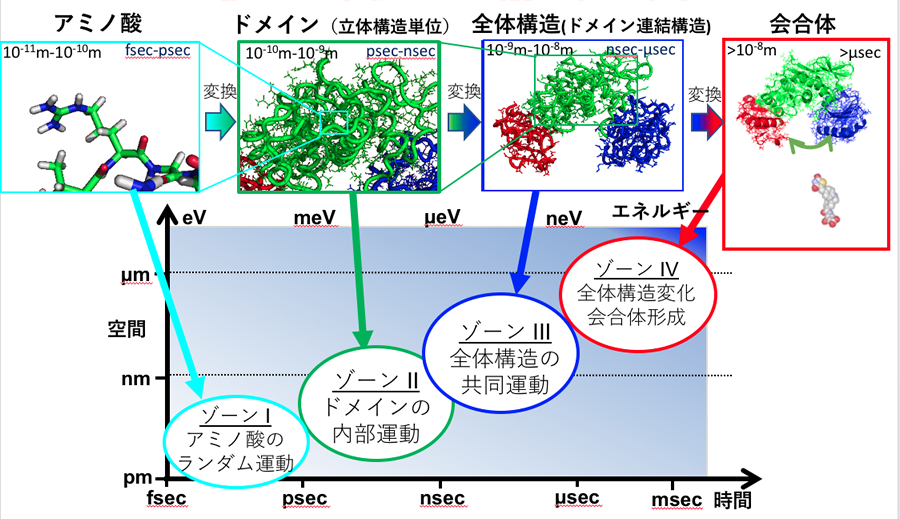
図1 杉山教授らのグループが主な研究対象としているのは、ゾーンⅡ(タンパク質のドメイン構造)、ゾーンⅢ(タンパク質の全体構造)とゾーンⅣ(タンパク質の会合体)である。
「アミノ酸からドメインを経て全体構造、さらには会合体に至るプロセスは、ほぼナノスケールでの現象です。だから我々は、タンパク質を『ナノマシン』と呼んでいます。そしてこのタンパク質は、まさにマシンと呼ぶにふさわしい特長を2つ備えています。ひとつはネジも歯車もないにも関わらず、複数のアミノ酸が、特定の形に凝集してマシンのように特定の機能を発現すること。もうひとつは、体内つまり生理活性状態におけるタンパク質は、マシンのように常に動き続けている点です」
溶液中で揺れ動くタンパク質の構造を解析するため、杉山教授は、量子ビーム(X線及び中性子)の散乱現象を利用して、構造解析を行う「小角散乱法」を採用した。
この方法で物質に量子ビームを当てると、ビームは物質内部で微細粒子に当たって散乱し、外に出てくる。従って、その散乱データを解析すると、微細粒子のサイズや分布、形状などが分かる。中性子を使えば0.1nmから1μmぐらいのスケールで、物質の内部構造を解析することができる。量子ビームとしてX線を使う場合は「SAXS(Small-Angle X-ray Scattering:X線小角散乱法)」、中性子を使う場合は「SANS(Small-Angle Neutron Scattering:中性子小角散乱法)」と呼ばれる。
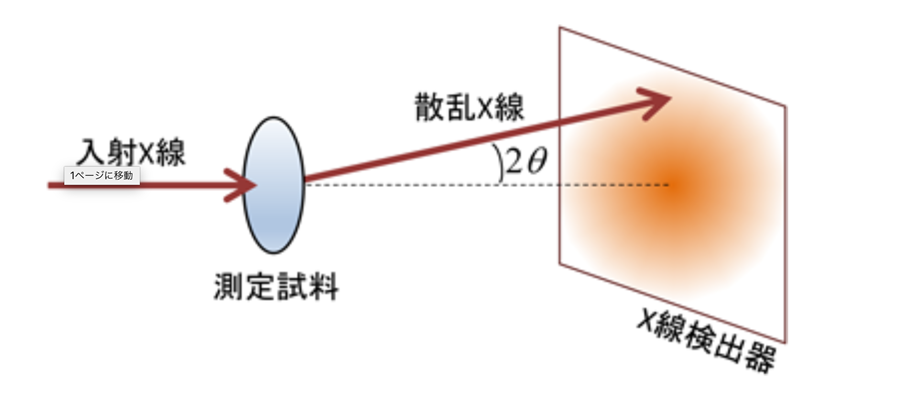
小角散乱法の基本原理。平行性の高い量子ビームを試料に当てると、試料を通過する際に内部の微細粒子によりビームが非常に小さい角度で散乱する。その結果を検出器で測定すれば、試料内部の構造を解析できる。
「小角散乱法」の特長は、次のように大きく5つ挙げられる。
1.0.1~1000nmと広範囲のスケールの構造を同一の測定原理に基づいて評価できる。
2.生体高分子や分子会合体のサイズや形状、それらの距離分布に関する情報を得られる。
3.固体、濃厚溶液、希薄溶液などさまざまな状態の物質をそのまま測定できる。
4.系全体の平均構造情報を評価できる。
5.非破壊での測定が可能。
この「小角散乱法」を使う利点を、杉山教授は次のように語る。
「同じように量子ビームを使う結晶解析法の場合、対象となるのはあくまでも結晶です。対してSAXSやSANSなら、試料が結晶でなくても解析できるので、溶液中や凝集体など非結晶状態のまま測定が可能です。つまり、実際の生体内部により近い条件で、タンパク質の挙動を測定できるのです」
さらに、中性子線を使うSANSの場合は、X線を使うSAXSでは解析できない要素まで解析可能だと杉山教授は語る。違いは色の濃淡である。
「X線でタンパク質を見ると、結果は一色で表示されます。ところが中性子を使うと、色の濃淡までをも見分けることができます。濃いところだけ、あるいは淡いところだけに注目して見ることも可能です。ということは、あらかじめ測定したい部分の濃淡を他の部分とは変えておけば、その動きをより精緻に見極められます」

研究室に設置されているX線小角散乱装置。主にソフトマターや生物試料のナノ構造解析に使用される。
このとき、タンパク質に“濃淡をつける”手段が重水素化である。測定したい試料タンパク質に含まれる水素原子を、重水素原子に置き換える手法だ。軽水素と重水素では、中性子を当てたときの散乱特性が異なるため、この手法を使えば、タンパク質の特定部分の構造に関する情報を得ることができる。
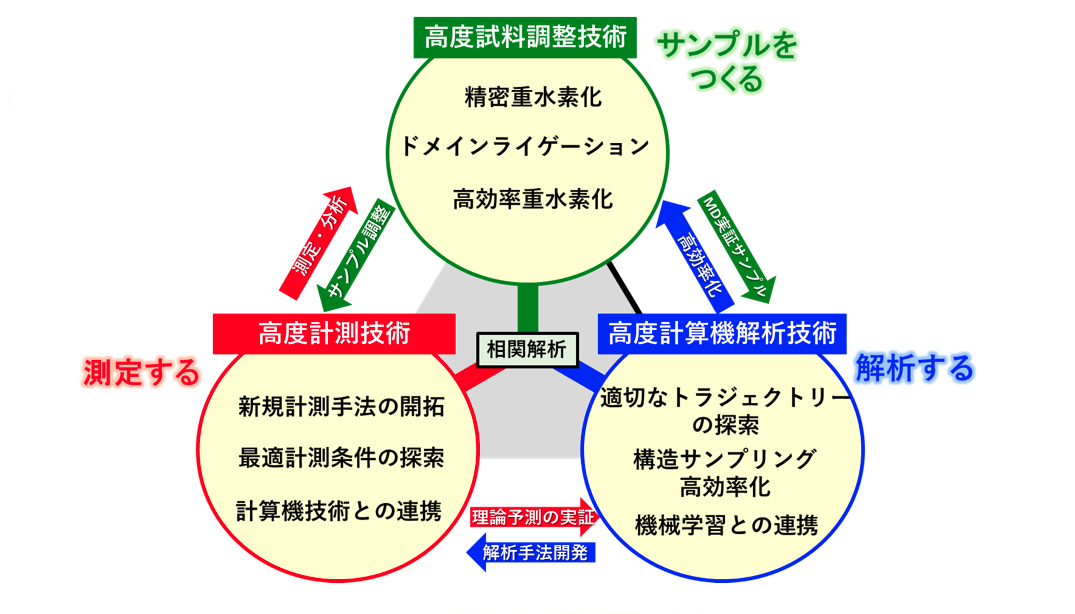
「重水素化してSANSを使えば、タンパク質を構成する複数のドメインのなかの特定ドメインだけを、選択的に細かく測定できます。さらに、得られたデータを、コンピュータを用いた分子動力学計算などで解析すると、実際にタンパク質が動いている様子をシミュレーションすることも可能です」
中性子小角散乱法による測定とコンピュータ解析。そしてもう一つ、測定を行う前の準備として研究に欠かせない技術がある。高度な試料調製技術である。

杉山研究室では、教授自らが研究員のデスクをまわり、研究の進捗について話し合いが行われる。研究員のいる部屋は、教授が命名したユニークな名前が付けられている。
タンパク質を測定するには、試料となるタンパク質が必要だ。タンパク質は大腸菌を培養してつくられるが、測定試料には「凝集」と呼ばれるタンパク質以外の成分も微量であるが含まれてしまう。タンパク質を精緻に測定するには、ゴミとなる凝集を可能な限り取り除かなければならない。そして、タンパク質の各ドメインを、重水素によって塗り分ける。ここまでのプロセスを経て、ようやく良質のサンプルができあがる。この試料を、X線や中性子を使って測定するわけだ。
「溶液中のタンパク質構造などを見るためには、可能な限り強いビーム光が望ましい。以前は、Spring8やKEKのような大型施設にある加速器から出てくる放射光を使う必要がありましたが、最近では研究室に設置可能なサイズのX線源でも強力なビームを出せるようになり、私の研究室にも先ほどの凝集を取り除く機器を装備したSAXS装置を備えていますし、小型ですがSANS装置もあります」
杉山教授の研究者としての出発点は、中性子にある。大学院時代に中性子の2次元検出器を自作し、これを使った中性子の小角散乱装置の開発研究で学位を取得した。その後、この装置を使って測定できる対象を広げていった結果、タンパク質と巡り合った。
ただし、タンパク質を精緻に中性子を用いて測定するには重水素化する必要がある。とはいえ重水素化したタンパク質は、当時は簡単にはつくれる代物ではなく、できたとしてもごく微量にとどまり、実験に必要な量をつくるには莫大なコストがかかるものだった。
「困っていた矢先に出会ったのが、分子科学研究所(大学共同利用機関法人)の加藤晃一教授でした。先生が『重水素化なら簡単にできるから、今度送るよ』と声をかけてくださり、逆に加藤先生は中性子小角散乱法をご存じなかったので、共同研究しましょうと話が進んで今に至ります」
高純度なサンプルをつくり、X線や中性子を使った測定を行う。得られたデータを、コンピュータを使って解析し、タンパク質の動きをシミュレーションによって可視化する。高度な試料調製技術と高度な計測技術、高度な計算機解析技術が三位一体となったとき、誰も見たことがないタンパク質の実像に迫れるのだ。
杉山教授らが研究対象として取り組んでいるテーマのひとつが、ヒトの眼の水晶体である。水晶体のタンパク質は、α、β、γと3種類の「クリスタリン」だけで構成されている。すなわち、溶液中で、これらのクリスタリン濃度を高めれば、眼球そのものとはいえないまでも、水晶体に近い状態を再現できるはずだ。
「水晶体では細胞が処分されません。そのため加齢に伴い少しずつ壊れていき、それが処分されずに残って白内障の発症に繋がります。一方で、αクリスタリンには、壊れた水晶体のタンパク質を治す作用があるらしいことが分かっていました。αクリスタリンは、タンパク質が25個から30個ぐらい集まった会合体です。ここで『30個ぐらい』と曖昧に表現したのにはれっきとした理由があります。というのも、この会合体は数が定まっていないからです。このことを詳しく調べてみると、会合体の間でタンパク質を交換していることが分かってきました。そしてこの交換作用がαクリスタリンのタンパク質修理機能に関連していると思っています」
水晶体内に近い環境におけるαクリスタリンの動態解析は、中性子小角散乱法を使わなければ実現しなかったものだ。杉山教授らの研究成果は、今後、白内障の治療に繋がる可能性をおおいに秘めている。
 杉山教授らのグループは、2020年6月に、小角散乱法により溶液中のタンパク質構造をより正確に評価する解析法を開発して発表した。X線や中性子を活用した小角散乱法では、あらかじめ可能な限り溶液中に含まれる凝集を取り除いておく必要があり、数%程度でも凝集が残ると、正確な散乱プロファイルを得られなくなる可能性があった。
杉山教授らのグループは、2020年6月に、小角散乱法により溶液中のタンパク質構造をより正確に評価する解析法を開発して発表した。X線や中性子を活用した小角散乱法では、あらかじめ可能な限り溶液中に含まれる凝集を取り除いておく必要があり、数%程度でも凝集が残ると、正確な散乱プロファイルを得られなくなる可能性があった。
「この問題を解消するために開発したのが、散乱プロファイルから凝集の影響を取り除く解析法『AUC-SAS』です。溶液を超遠心分析(AUC)にかけた結果を元に、凝集の種類と濃度に関するデータを収集し、散乱プロファイルから凝集の影響を取り除くのです。AUC-SASを使えば、今後の見通しとして、生体により近い環境である複雑な多成分溶液中で、狙ったタンパク質の構造を解析できる可能性があります」
タンパク質は、構成するアミノ酸の種類や数、結合の順序などによって種類が異なる。分子量でみれば、約4000前後から最大では数千万からなるタンパク質もある。しかも複数のタンパク質が結合した会合体まであり、それぞれに果たす機能が異なってくる。
「タンパク質の研究テーマは、正直なところ尽きることがありません。マルチドメインタンパク質の溶液中の構造を例とするなら、水晶体を構成するβB2クリスタリンは、2つのドメインを持つことが明らかになっています。これを結晶構造解析するとドメイン間の距離が44Å(オングストローム、1Å=0.1ナノメートル(nm))と出ます。ただし、結晶化されたタンパク質は、細胞内での実態とは異なります。溶液中の距離を我々が解析した結果、2つのドメインは32Åまで近接していることが分かりました。ここまで近接しているから必要な機能を発現するのです」
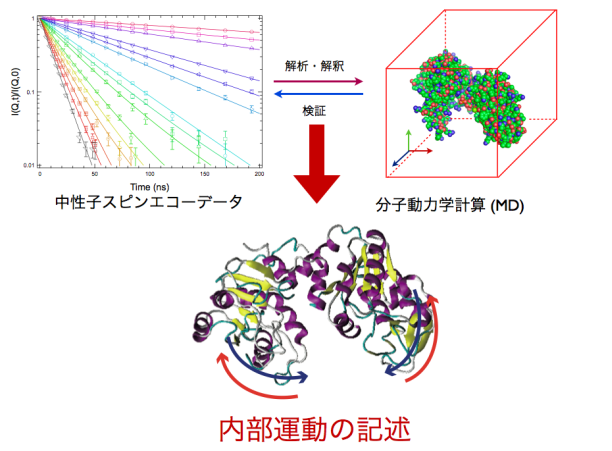
杉山研究室では、測定データを分子動力学計算により解析する手法の確立に取り組んでいる。その最終ゴールは、タンパク質の内部運動の記述、すなわち「見える化」である。
マルチドメインタンパク質では、会合体内部での各ドメインの働きが、タンパク質全体として発現する機能を左右する。個々の機能を解明するには、コンピュータを駆使して、高精度な測定データを解析する必要がある。データに基づくシミュレーション精度が高まれば、生体内でのタンパク質の動きをリアルに再現できる。
「今後の課題はいくらでもあるというか、この研究に終わりはないというのが正直なところです。何しろ相手が10万種類以上あるとされるタンパク質だけに、何か一つ研究すれば、そこから芋づる式に次のテーマが次々と出てくるわけです。とはいえ当面のゴールは、重水素化したタンパク質を使い、生体内部に限りなく近い環境で、狙い定めたドメインをクリアに見ること、そして、その技術を汎用化することです。さらにその先の夢想に近いテーマを語るなら、思い通りに動くタンパク質をつくってみたい。そんなタンパク質をいくつかつくって、どれが一番優れているかを競わせてみたいですね」
杉山教授は小学生の頃から、科学者以外に自分の将来の姿はないと覚悟を決めていた。そして「ノーベル賞を取りたいから」と京都大学理学部へ進学した。ところが、そこで大きな挫折を味わった。
「数学科を筆頭に、理学部には飛び抜けて頭の良い面々がいくらでもいました。私にはさっぱりわからない、極めて難解な数学の問題なども、数式で考える以前にビジュアルとして答えが見えているような連中です。それでも何とか志望していた物理系の教室に進み、そこから中性子の研究を始めました。大きな転機となったのは、加藤先生と重水素化に出会ったことでしょう。この出会いをきっかけに、タンパク質の研究に本格的に取り組むようになり、将来の道が開けた気分になりました」
杉山教授の飾らない人柄に惹かれるようにして、研究室には優れた人材が集まっている。教授は一人ひとりに、その専門技術にちなんだ肩書きをつけている。新規の試料作製技術の開発を担当するポスドクには「破砕の(タンパク質をつくる時に大腸菌を大量に破砕するので)」Aさん、分子動力学計算で解析を担当するのは「阻止禍の(粗視化をもじったもの)」Bさんといった案配だ。いわゆる“あだ名”だが、それを研究者たちもどことなく喜んでいるように思える。
ユーモアあふれる杉山教授が、本当に思い通りに動くタンパク質をつくり出す日は、それほど先の未来のことではないはずだ。


原子炉による実験及びこれに関連する研究を行う研究機関。1963年に京都大学原子炉実験所として設立され、原子力と放射線の利用に関する研究教育活動に取り組んでいる。京都大学研究用原子炉と京都大学臨海集合体実験装置の2つの原子炉を備えている。研究分野は幅広く、原子力基礎科学から粒子線物質科学、放射線生命医科学などに及ぶ。2018年より現在の名称に変更された。
【取材・文:竹林篤実 撮影:大島拓也】




 bana1.png (300px×80px)
bana1.png (300px×80px) bana1_e.png (300px×80px)
bana1_e.png (300px×80px) NovelPrize2015.png
NovelPrize2015.png